فیزیولوژی دانشی است که وظیفهاش بررسی عملکرد (Function) موجودات زنده است. ماهیت بررسی در این علم ، وظیفه و کارکرد اندامهاست. نام قدیمی فیزیولوژی وظایفالاعضا بوده است. فیزیولوژی گیاهی ، مطالعه اعمال حیاتی گیاه ، فرایندهای رشد و نمو ، متابولیزم و تولید مثل گیاهان است.
دید کلی
کشف قوانینی که بر تغذیه گیاه و رشد و نمو آن حکومت میکند، شناخت توانایی واقعی سلولها در انجام فعالیتهای بیولوژیک و همچنین ارائه روشهایی که ظهور یکی از توانائیهای سلولی را امکانپذیر میسازد، هدف اساسی فیزیولوژی گیاهی محسوب میشود. همانطور که مسیر روشن بسیاری از اکتشافات نظری ، منشا پیشرفتهایی در یکی از شاخههای تجربی علوم است، نتایج حاصل از مطالعاتی که در همه شئون علمی بالاخص در فیزیولوژی گیاهی صورت گرفته، باعث توسعه و پیشرفت واقعی کشاورزی شده و آن را از صورت ابتدایی خود در نخستین روزهای ظهور انسان به صورت کاملا پیشرفته امروزی ، مبدل ساخته است.
از طرف دیگر ، ترقیات سریع فیزیولوژی گیاهی نیز خود مدیون ترقیات علوم دیگری مانند فیزیک و شیمی است، زیرا عملا کلیه اعمال متابولیزم سلولها بر اساس قوانینی تفسیر میشوند که در مورد عالم بیجان شناخته شدهاند. شک نیست که علم فیزیولوژی گیاهی ، علمی است تجربی و همه کوششهایی که در این زمینه صورت میگیرند، به شناسایی بیش از پیش ماده زنده منجر میشوند. به علاوه فیزیولوژی گیاهی ، علم پایه مستقلی است که دارای مفاهیم خاصی بوده، شیوه مخصوصی در تجربیات آن مشاهده میشود.
موضوعات مطرح شده در فیزیولوژی گیاهی
فیزیولوژی گیاهی را میتوان مطالعه اعمال حیاتی گیاه ، فرایندهای چرخهای متحرک رشد ، متابولیزم و تولید مثل دانست. مباحث زیادی در فیزیولوژی گیاهی بحث میشود و در هیچ علمی ، نحوه پیشرفت واضحتر از زمینه فیزیولوژی گیاهی نیست. از مباحثی که در فیزیولوژی گیاهی بحث میشود، میتوان به موارد زیر اشاره کرد.
تغذیه و جذب در گیاهان
انجام صحیح فرایندهای متابولیزمی مستلزم وجود عناصری است که باید به صورت اکسید شده یا احیا شده ، معدنی و یا آلی جذب سلولها شده، احتیاجات آنها را از نظر ماده و انرژی تامین کنند. مقدار و نوع این احتیاجات تابعی از شدت و نوع واکنشهای متابولیزمی بوده و به همین مناسبت هر موجودی از نظر قدرت سنتز و طریقه تحصیل انرژی با موجود دیگر متفاوت است.
موجودات زنده را از نظر قدرت سنتز و همانند سازی به دو دسته اتوتروف و هتروتروف تقسیم می کنند. موجودات اتوتروف موجوداتی را گویند که از ترکیبات سادهای نظیر دیاکسید کربن و ترکیبات معدنی مختلف مانند نیتروژن معدنی ، میتوانند کلیه احتیاجات خود را برطرف سازند که گیاهان در این گروه قرار میگیرند.
احتیاجات گیاهان نسبت به انرژی
سلولهای گیاهی انرژی موجود در مواد تشکیل دهنده خود را به صور مختلف زیر از دست میدهند.
به صورت انرژی حرارتی که در بعضی موارد مانند گل آذین گل شیپوری کاملا آشکار است.
به صورت انرژی نورانی مانند فلورسانس کلروفیل
به صورت انرژی مکانیکی مانند سیکلوز در سیتوپلاسم
به صورت انرژی الکتریکی که نتیجه آن برقراری اختلاف پتانسیل بین اعضای مختلف گیاهان است.
احتیاجات گیاهان نسبت به مواد
میزان این احتیاجات در نمونههای مختلف گیاهی ، متفاوت است. رفع احتیاجات یک گیاه بالغ در درجه اول به منظور جبران موادی است که این گیاه در طول حیات از دست میدهد. در درجه دوم ، رشد و نمو یک گیاه احتیاجات احتمالی دیگری بوجود میآورد. کلیه این احتیاجات بوسیله منابع طبیعی مختلفی تامین میشوند که عبارتند از: خاک ، هوا ، آب و محیطهای آلی.
بطور کلی در بخش تغذیه و جذب مباحث مختلفی بحث میشود: احتیاجات گیاهان ، نقش عمومی و اختصاصی عناصر و علائم کمبودهای آنها ، محلولهای غذایی و کودهای شیمیایی ، تغذیه نیتروژن معدنی و آلی ، چرخه متابولیزمی نیتروژن ، گوگرد و فسفر ، رابطه آب و خاک ، گردش مواد در گیاه ، جذب مواد معدنی ، مکانیزم جذب مواد و ... .
فتوسنتز
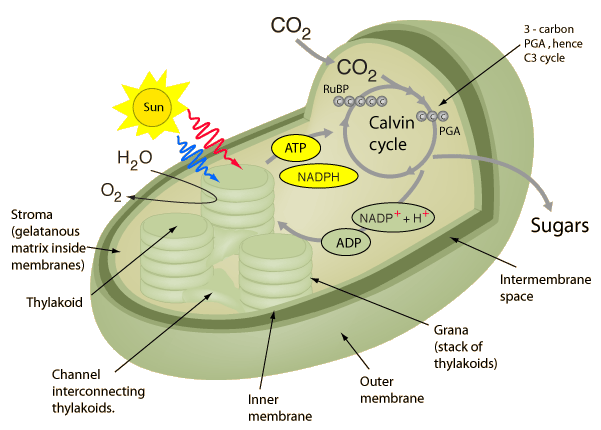
زندگی در روی کره زمین به انرژی حاصل از خورشید وابسته است. فتوسنتز از نظر لغوی به معنی تولید با استفاده از نور خورشید است. در فتوسنتز ، انرژی خورشیدی برای اکسید کردن آب ، آزاد شدن اکسیژن و نیز احیا کردن به ترکیبات آلی و در نهایت قند بکار میرود. فتوسنتز شامل دو دسته از واکنشهاست: واکنشهای نوری و واکنشهای تاریکی.
بطور کلی در بخش فتوسنتز مباحث مختلفی بحث می شود:
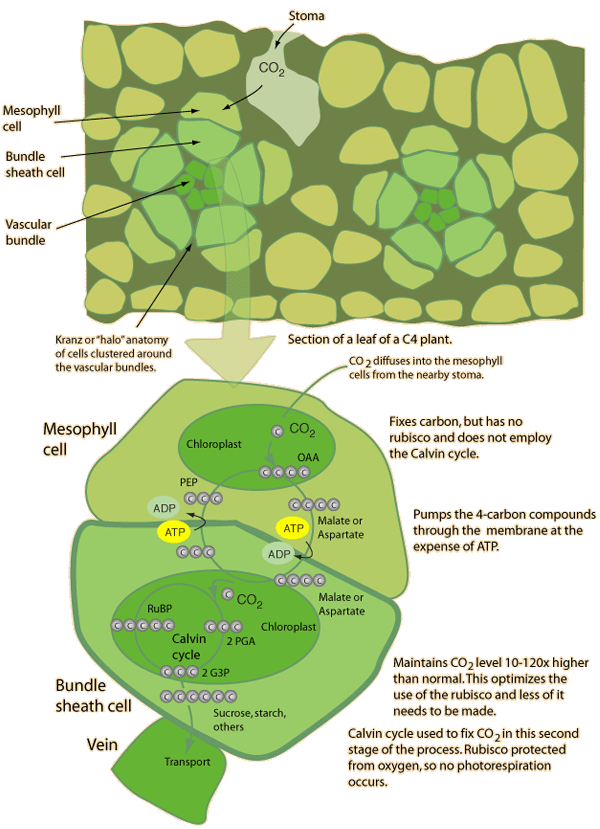
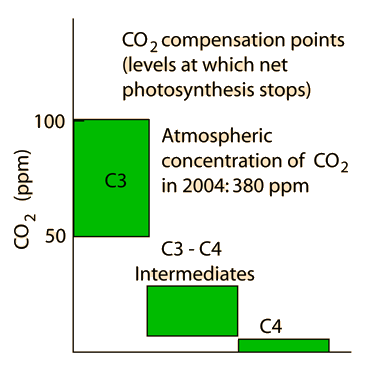
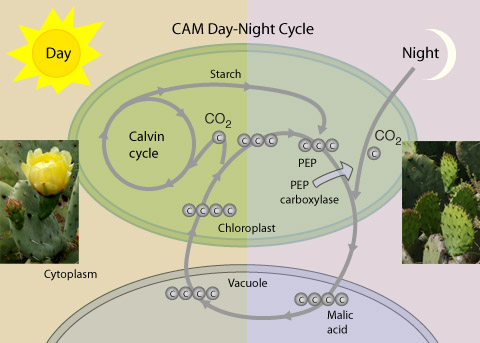
مفاهیم کلی در مورد فتوسنتز ، عملکرد کوانتومی نور ، ساختمان دستگاه فتوسنتزی ، ساختار تیلاکوئیدها در کلروپلاست ، گیرندههای نوری ، فتوسیستمهای I و II ، مکانیزم انتقال الکترون و پروتون در کلروپلاستها ، ژنوم کلروپلاست ، چرخه احیای فتوسنتزی ، تنفس نوری ، چرخه احیای فتوسنتزی ، چرخه احیای کربن در گیاهان CAM(کراسولاسه) ، سنتز نشاسته و ساکارز در گیاهان و ... .
تنفس
تنفس فرایندی است که انرژی ذخیره شده در مواد انرژیزا مانند کربوهیدراتها را به شیوهای کنترل شده ، آزاد میکند. در طی تنفس انرژی آزاد ، رها شده و به شکل ATP در میآید که این شکل از انرژی میتواند به سهولت برای نگهداری و رشد گیاه مورد استفاده قرار گیرد.
مباحثی که در مورد تنفس در فیزیولوژی گیاهی ، بحث میشود، به صورت زیر است:
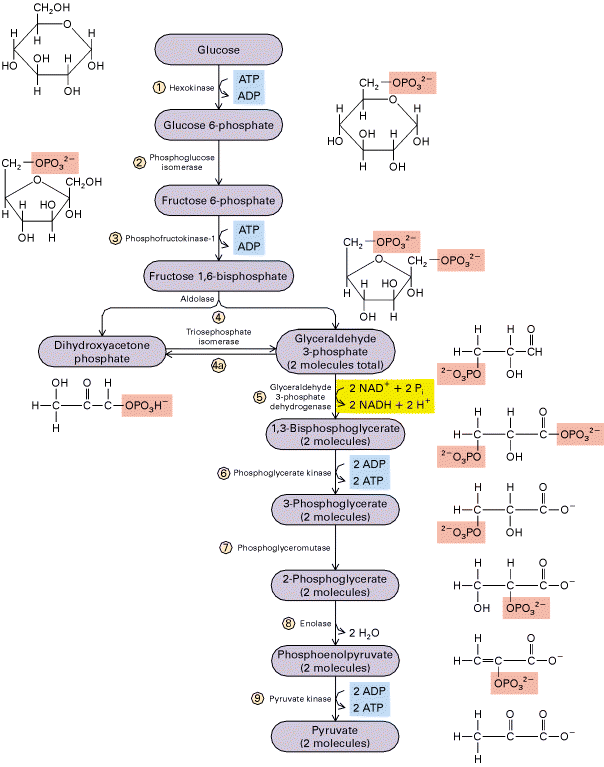
تنفس هوازی و بیهوازی ، ساختمان میتوکندریها ، گلیکولیز و چرخه کربس ، زنجیره انتقال الکترون در میتوکندری ، مسیر پنتوز فسفات و ... .
رشد و نمو گیاهی
رشد و نمو اساسا از پدیدههای مهم در طی انتوژنی گیاه است. رشد و نمو تحت تاثیر عوامل متعدد محیطی و ژنتیکی قرار دارد. البته عامل مهم تعیین کننده الگوهای رشد و نمو ، عمدتا پایگاه ژنتیکی دارد. رشد عبارت است تغییرات کمی و افزایش غیر قابل برگشت در ابعاد یک موجود یا یک اندام. به مجموعه تغییراتی که ماهیت کیفی دارند، به اضافه تغییرات کمی (رشد) ، نمو اطلاق میشود.
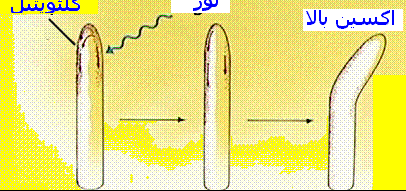
مباحثی که در رشد و نمو گیاهی بحث میشود، به صورت زیر است. سینتیک رشد ، تروپیسمها یا گرایشها در گیاهان ، جنبشهای گیاهان ، تنظیم کنندهها یا هورمونهای رشد در گیاه مانند اکسین ، جیبرلین و ... ، مکانیزم تشکیل گل و فتوپریودیسم ، فیتوکرومها و دیگر پذیرندههای نوری و ... .
ارتباط فیزیولوژی گیاهی با سایر علوم
فیزیولوژی گیاهی با بسیاری از علوم ، ارتباط دارد. مانند بیوشیمی ، بیوفیزیک و بیولوژی مولکولی. البته فیزیولوژیستها مکررا از نتایج تحقیقات بیوشیمیستها و متخصصان بیوفیزیک و بیولوژی مولکولی استفاده میکنند و متقابلا دانشمندان رشتههای دیگر نیز از نتایج آزمایشات فیزیولوژی گیاهی ، بهرهمند میشوند.
در حقیقت این رشتههای مرتبط ، با هم یک مجموعه ایجاد میکنند و مرزهای تعریف شده عمدتا مصنوعی هستند. بنابراین آشنایی با مبانی بیوفیزیک ، بیوشیمی و بیولوژی مولکولی ، غیرقابل تفکیک با فیزیولوژی گیاهی هستند.
دید کلی
کشف قوانینی که بر تغذیه گیاه و رشد و نمو آن حکومت میکند، شناخت توانایی واقعی سلولها در انجام فعالیتهای بیولوژیک و همچنین ارائه روشهایی که ظهور یکی از توانائیهای سلولی را امکانپذیر میسازد، هدف اساسی فیزیولوژی گیاهی محسوب میشود. همانطور که مسیر روشن بسیاری از اکتشافات نظری ، منشا پیشرفتهایی در یکی از شاخههای تجربی علوم است، نتایج حاصل از مطالعاتی که در همه شئون علمی بالاخص در فیزیولوژی گیاهی صورت گرفته، باعث توسعه و پیشرفت واقعی کشاورزی شده و آن را از صورت ابتدایی خود در نخستین روزهای ظهور انسان به صورت کاملا پیشرفته امروزی ، مبدل ساخته است.
از طرف دیگر ، ترقیات سریع فیزیولوژی گیاهی نیز خود مدیون ترقیات علوم دیگری مانند فیزیک و شیمی است، زیرا عملا کلیه اعمال متابولیزم سلولها بر اساس قوانینی تفسیر میشوند که در مورد عالم بیجان شناخته شدهاند. شک نیست که علم فیزیولوژی گیاهی ، علمی است تجربی و همه کوششهایی که در این زمینه صورت میگیرند، به شناسایی بیش از پیش ماده زنده منجر میشوند. به علاوه فیزیولوژی گیاهی ، علم پایه مستقلی است که دارای مفاهیم خاصی بوده، شیوه مخصوصی در تجربیات آن مشاهده میشود.
موضوعات مطرح شده در فیزیولوژی گیاهی
فیزیولوژی گیاهی را میتوان مطالعه اعمال حیاتی گیاه ، فرایندهای چرخهای متحرک رشد ، متابولیزم و تولید مثل دانست. مباحث زیادی در فیزیولوژی گیاهی بحث میشود و در هیچ علمی ، نحوه پیشرفت واضحتر از زمینه فیزیولوژی گیاهی نیست. از مباحثی که در فیزیولوژی گیاهی بحث میشود، میتوان به موارد زیر اشاره کرد.
تغذیه و جذب در گیاهان
انجام صحیح فرایندهای متابولیزمی مستلزم وجود عناصری است که باید به صورت اکسید شده یا احیا شده ، معدنی و یا آلی جذب سلولها شده، احتیاجات آنها را از نظر ماده و انرژی تامین کنند. مقدار و نوع این احتیاجات تابعی از شدت و نوع واکنشهای متابولیزمی بوده و به همین مناسبت هر موجودی از نظر قدرت سنتز و طریقه تحصیل انرژی با موجود دیگر متفاوت است.
موجودات زنده را از نظر قدرت سنتز و همانند سازی به دو دسته اتوتروف و هتروتروف تقسیم می کنند. موجودات اتوتروف موجوداتی را گویند که از ترکیبات سادهای نظیر دیاکسید کربن و ترکیبات معدنی مختلف مانند نیتروژن معدنی ، میتوانند کلیه احتیاجات خود را برطرف سازند که گیاهان در این گروه قرار میگیرند.
احتیاجات گیاهان نسبت به انرژی
سلولهای گیاهی انرژی موجود در مواد تشکیل دهنده خود را به صور مختلف زیر از دست میدهند.
به صورت انرژی حرارتی که در بعضی موارد مانند گل آذین گل شیپوری کاملا آشکار است.
به صورت انرژی نورانی مانند فلورسانس کلروفیل
به صورت انرژی مکانیکی مانند سیکلوز در سیتوپلاسم
به صورت انرژی الکتریکی که نتیجه آن برقراری اختلاف پتانسیل بین اعضای مختلف گیاهان است.
احتیاجات گیاهان نسبت به مواد
میزان این احتیاجات در نمونههای مختلف گیاهی ، متفاوت است. رفع احتیاجات یک گیاه بالغ در درجه اول به منظور جبران موادی است که این گیاه در طول حیات از دست میدهد. در درجه دوم ، رشد و نمو یک گیاه احتیاجات احتمالی دیگری بوجود میآورد. کلیه این احتیاجات بوسیله منابع طبیعی مختلفی تامین میشوند که عبارتند از: خاک ، هوا ، آب و محیطهای آلی.
بطور کلی در بخش تغذیه و جذب مباحث مختلفی بحث میشود: احتیاجات گیاهان ، نقش عمومی و اختصاصی عناصر و علائم کمبودهای آنها ، محلولهای غذایی و کودهای شیمیایی ، تغذیه نیتروژن معدنی و آلی ، چرخه متابولیزمی نیتروژن ، گوگرد و فسفر ، رابطه آب و خاک ، گردش مواد در گیاه ، جذب مواد معدنی ، مکانیزم جذب مواد و ... .
فتوسنتز
زندگی در روی کره زمین به انرژی حاصل از خورشید وابسته است. فتوسنتز از نظر لغوی به معنی تولید با استفاده از نور خورشید است. در فتوسنتز ، انرژی خورشیدی برای اکسید کردن آب ، آزاد شدن اکسیژن و نیز احیا کردن به ترکیبات آلی و در نهایت قند بکار میرود. فتوسنتز شامل دو دسته از واکنشهاست: واکنشهای نوری و واکنشهای تاریکی.
بطور کلی در بخش فتوسنتز مباحث مختلفی بحث می شود:
مفاهیم کلی در مورد فتوسنتز ، عملکرد کوانتومی نور ، ساختمان دستگاه فتوسنتزی ، ساختار تیلاکوئیدها در کلروپلاست ، گیرندههای نوری ، فتوسیستمهای I و II ، مکانیزم انتقال الکترون و پروتون در کلروپلاستها ، ژنوم کلروپلاست ، چرخه احیای فتوسنتزی ، تنفس نوری ، چرخه احیای فتوسنتزی ، چرخه احیای کربن در گیاهان CAM(کراسولاسه) ، سنتز نشاسته و ساکارز در گیاهان و ... .
تنفس
تنفس فرایندی است که انرژی ذخیره شده در مواد انرژیزا مانند کربوهیدراتها را به شیوهای کنترل شده ، آزاد میکند. در طی تنفس انرژی آزاد ، رها شده و به شکل ATP در میآید که این شکل از انرژی میتواند به سهولت برای نگهداری و رشد گیاه مورد استفاده قرار گیرد.
مباحثی که در مورد تنفس در فیزیولوژی گیاهی ، بحث میشود، به صورت زیر است:
تنفس هوازی و بیهوازی ، ساختمان میتوکندریها ، گلیکولیز و چرخه کربس ، زنجیره انتقال الکترون در میتوکندری ، مسیر پنتوز فسفات و ... .
رشد و نمو گیاهی
رشد و نمو اساسا از پدیدههای مهم در طی انتوژنی گیاه است. رشد و نمو تحت تاثیر عوامل متعدد محیطی و ژنتیکی قرار دارد. البته عامل مهم تعیین کننده الگوهای رشد و نمو ، عمدتا پایگاه ژنتیکی دارد. رشد عبارت است تغییرات کمی و افزایش غیر قابل برگشت در ابعاد یک موجود یا یک اندام. به مجموعه تغییراتی که ماهیت کیفی دارند، به اضافه تغییرات کمی (رشد) ، نمو اطلاق میشود.
مباحثی که در رشد و نمو گیاهی بحث میشود، به صورت زیر است. سینتیک رشد ، تروپیسمها یا گرایشها در گیاهان ، جنبشهای گیاهان ، تنظیم کنندهها یا هورمونهای رشد در گیاه مانند اکسین ، جیبرلین و ... ، مکانیزم تشکیل گل و فتوپریودیسم ، فیتوکرومها و دیگر پذیرندههای نوری و ... .
ارتباط فیزیولوژی گیاهی با سایر علوم
فیزیولوژی گیاهی با بسیاری از علوم ، ارتباط دارد. مانند بیوشیمی ، بیوفیزیک و بیولوژی مولکولی. البته فیزیولوژیستها مکررا از نتایج تحقیقات بیوشیمیستها و متخصصان بیوفیزیک و بیولوژی مولکولی استفاده میکنند و متقابلا دانشمندان رشتههای دیگر نیز از نتایج آزمایشات فیزیولوژی گیاهی ، بهرهمند میشوند.
در حقیقت این رشتههای مرتبط ، با هم یک مجموعه ایجاد میکنند و مرزهای تعریف شده عمدتا مصنوعی هستند. بنابراین آشنایی با مبانی بیوفیزیک ، بیوشیمی و بیولوژی مولکولی ، غیرقابل تفکیک با فیزیولوژی گیاهی هستند.